Protein turnover within a cell is a normal process that is highly regulated. Degradation destined proteins are degraded via distinct pathways.
For example, a newly synthesized, yet misfolded protein is degraded via the UPS or ERAD pathways in ubiquitin dependent manner. In contrast, protein aggregates or oxidatively modified proteins are predominantly degraded via autophagy.
However it is now clear that these pathways while complementary may serve overlapping functions. Our research is particularly interested in the communication between autophagy and the UPS.
Ubiquitin Proteasome System (UPS)
A degradation destined protein (e.g. misfolded, short-lived or regulated by its degradation) are tagged with ubiquitin by a specific E3 ubiquitin ligase. This single ubiquitin is then extended into a polyubiquitin chain which targets the protein for degradation by the proteasome. Several proteins regulate this process including E3 ligases, deubiquitinating enzymes, proteasome targeting proteins and the 26S proteasome itself.

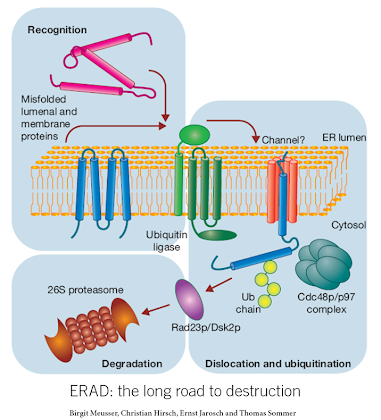
Endoplasmic reticulum associated degradation (ERAD)
ERAD is a specialized type of protein degradation which is specific to proteins within the secretory pathway (e.g. ER lumen, secreted or transmembrane proteins). In order for an ER synthesized protein to be degraded, it must (1) be recognized as misfolded, (2) be extruded from the ER through a channel known as the retrotranslocon, and (3) once in the cytosol (where protein degradation machinery exists) it is ubiquitinated and targeted for the proteasome. ERAD requires several additional proteins in addition to an intact UPS for its function. These include ER chaperones, ER localized E3 ligases and the retrotranslocon.
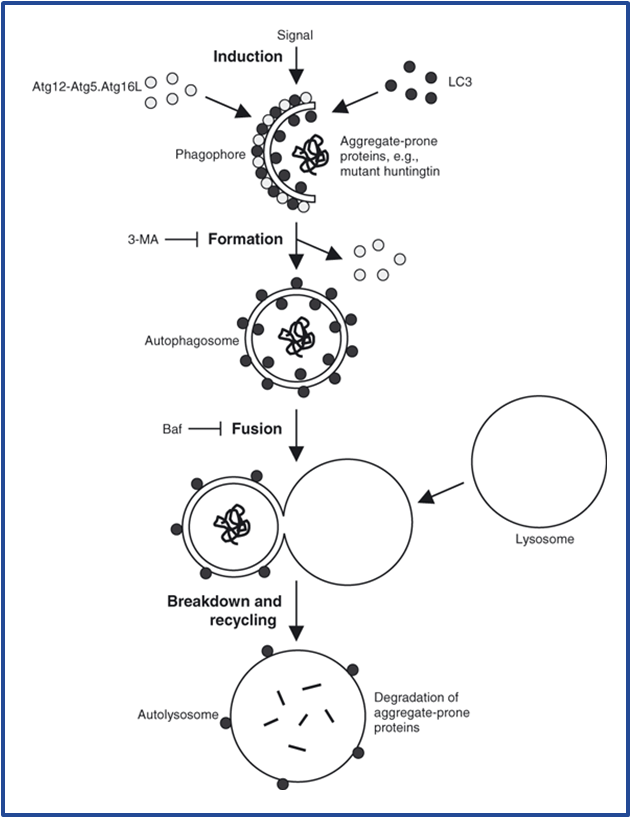
Autophagy
Autophagy, specifically macroautophagy, performs bulk degradation of protein and organelles in a cell. It occurs when a signal (either extracellular or intracellular) induces the formation of a phagophore. A phagophore begins as a preautophagic structure (vesicle) that extends in size and eventually engulfs a region of cytoplasm. Once the ends of a phagophore fuse it is termed an autophagosome. Autophagosomes are double membrane lined vesicles that contain degradation destined proteins and organelles. They are easily recognized by fluorescent microscopy because the contain the protein ATG8/LC3. An autophagosome then fuses with an acidic lysosome generating and an autolysome which degrades the contents of the autophagosome.
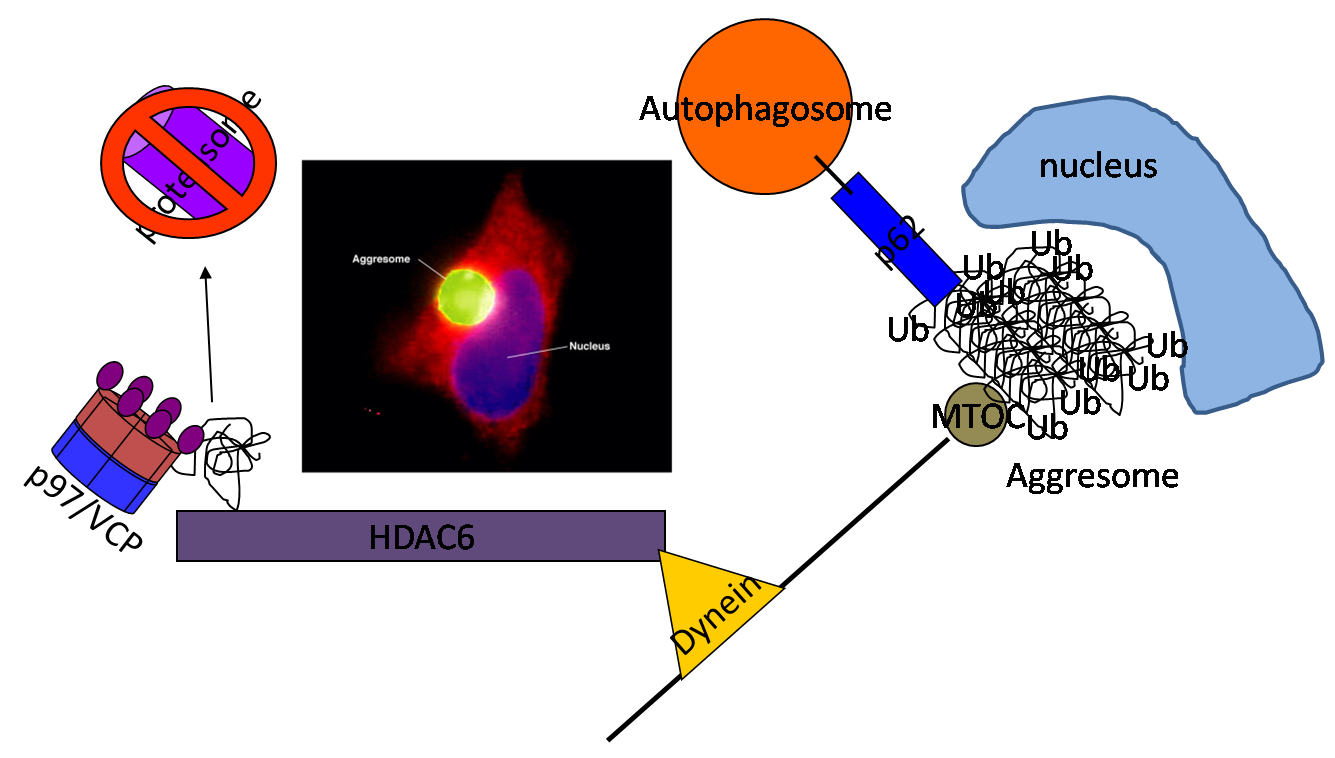
Aggresomes and inclusion bodies
Aggresomes are microtubule dependent perinuclear accumulations of ubiquitinated protein aggregates in a cell. Aggresomes form when the proteasome is inhibited or when a misfolded or aggregated protein is unable to be degraded via the UPS. Aggresomes are similar to inclusion bodies seen in degenerative diseases. Aggresomes sequester toxic protein aggregates and allow them to be degraded via autophagy. Several proteins including VCP are essential for aggresome formation. One such protein HDAC6 can facilitate the autophagic degradation of ubiquitinated proteins and protein aggregates in the setting of proteasome impairment. This enhanced autophagic clearance of protein inclusions is neuroprotective in neurodegenerative disease models.